una cascata di eventi divinamente i sincronizzati
L’ipertrofia muscolare è un evento determinato da diversi fattori.
A determinare tale evento è lo stress meccanico (tensione) a cui sono sottoposte le cellule muscolari ed il conseguente danno che scompensa temporaneamente l’architettura di queste cellule ed ancora il cambiamento degli equilibri chimici intracellulari che si verifica durante e dopo lo stress.
Il danno muscolare sembra essere direttamente proporzionale all’intensità relativa, ovvero alla percentuale del cedimento muscolare. Più una serie allenante si avvicina al cedimento e più questo viene tradotto in danno in sede miocitaria.
Ovviamente è fondamentale l’utilizzo di un quantitativo di resistenza (carico) sufficiente a garantire il reclutamento ottimale, attraverso meccanismi descritti in seguito.
Significa che il cedimento oltre le dieci ripetizioni serve a ben poco…
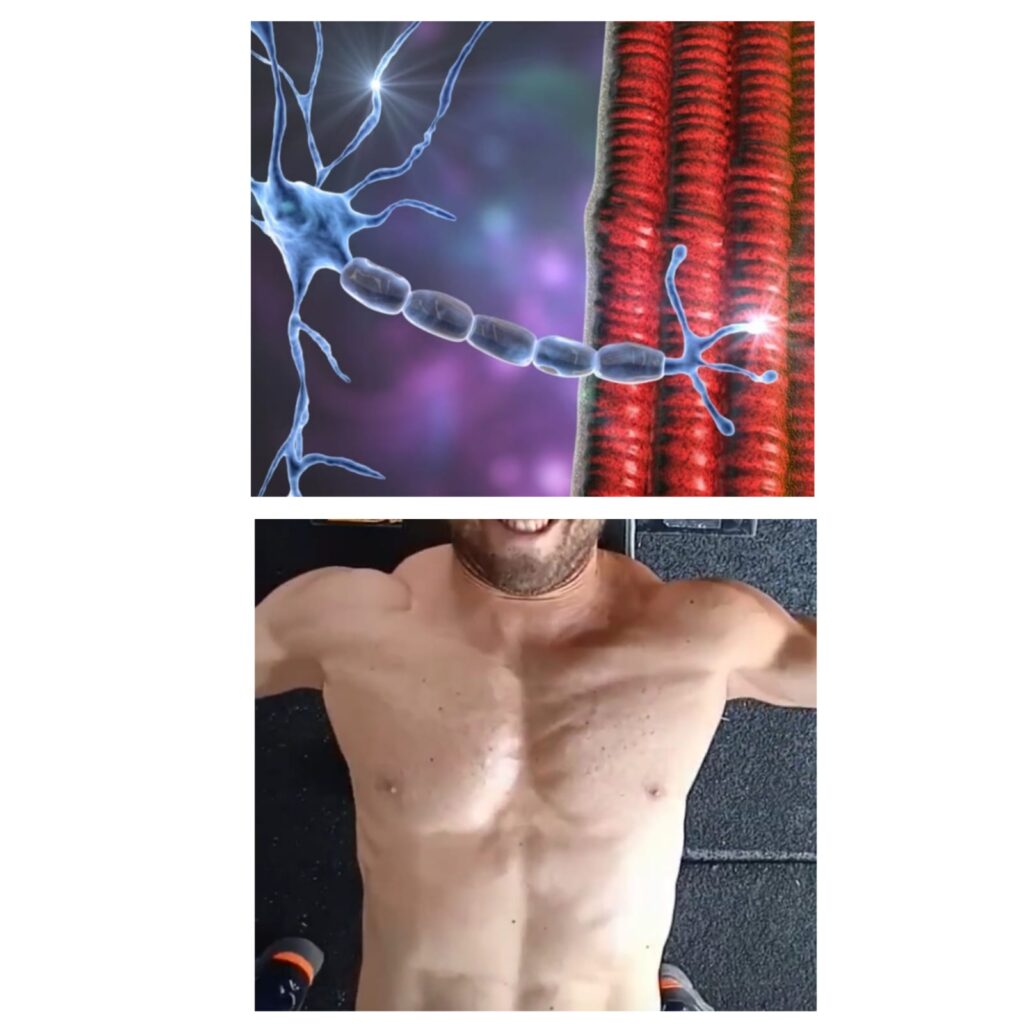
La contrazione muscolare rappresenta l’ultima fase di una concatenazione di fenomeni che partono dal sistema nervoso coinvolgendo cellule nervose, mediatori chimici, elettroliti e membrane.
STRUTTURA E FUNZIONE DELLE CELLULE NERVOSE
I neuroni motori o motoneuroni avviano il fenomeno della contrazione muscolare attraverso un impulso nervoso, che consiste in un rapido cambiamento del potenziale elettrico, chiamato potenziale d’azione.
I motoneuroni sono localizzati nel sistema nervoso centrale e periferico, nello specifico a livello del telencefalo (corteccia cerebrale) e del midollo spinale.
Il motoneurone è una cellula, che si compone di un nucleo, un corpo, delle ramificazioni in grado di ricevere stimoli periferici di natura eccitatoria o inibitoria e di un prolungamento, l’assone, un canale lungo il quale si propagano gli stimoli efferenti (significa dal neurone verso gli effettori, in questo caso i muscoli).
Gli assoni sono raggruppati in fasci circondati da tessuto connettivo a formare i processi nervosi.
Le cellule nervose sono circondate da tessuti non nervosi come la mielina, una guaina che circonda gli assoni assicurandogli protezione, le cellule di Schwann responsabili della sintesi della mielina e gli astrociti che apportano nutrienti al neurone e ne regolano l’equilibrio elettrolitico.
Il fluido che si trova nello spazio interstiziale tra le membrane dei neuroni, ha un’elevata concentrazione di ioni sodio e anioni cloro, mentre all’interno della membrana è alta la concentrazione di ioni potassio ed altri grandi anioni.
La concentrazione di sodio all’esterno è dieci volte superiore rispetto all’ interno. Viceversa quella del potassio è venticinque volte superiore all’ interno rispetto all’ esterno.
Questo fattore è di fondamentale importanza, in quanto rende la cellula nervosa negativa all’interno e positiva all’esterno.
I canali presenti sulla membrana sono permeabili al potassio, che potrebbe diffondere all’esterno, mentre quelli del sodio sono chiusi. Essi infatti si aprono solo quando ricevono il passaggio del potenziale d’azione.
In ogni caso il potassio non esce dall’assone perché respinto dalle cariche positive (stesso segno) del sodio, ed allo stesso tempo è attirato dai grandi anioni all’interno, che a loro volta non escono perché troppo grandi per attraversare qualsiasi canale di membrana.
Al passaggio del potenziale d’azione si aprono i canali del sodio, che entra nell’assone cambiando il voltaggio da negativo a positivo. Come effetto di bilanciamento parte degli ioni potassio fuoriescono, per poi rientrare spinti dagli astrociti.
Il sodio è invece rimosso dall’interno della cellula grazie a delle pompe che consumano ATP.
Questo fenomeno si verifica in un millisecondo e solo un milionesimo del sodio presente all’esterno attraversa la membrana in questo lasso di tempo infinitesimale.
PROPAGAZIONE DELL’ IMPULSO.
LA GIUNZIONE NEUROMUSCOLARE
Ogni singola fibra muscolare è circondata dal sarcolemma, che funge da membrana della cellula muscolare.
Sotto il sarcolemma è presente un sistema reticolare, il reticolo sarcoplasmatico, che a sua volta è affiancato da canali paralleli alle unità contrattili delle fibre, che si intersecano in prossimità dei centri dei sarcomeri formando delle cisterne in cui sono contenuti gli ioni calcio.
Le cisterne confinano (ma non comunicano) con formazioni tubulari detti Tubuli T, invaginazioni del sarcolemma che decorrono trasversalmente alle unità contratti e servono a trasmettere il potenziale d’azione alle cisterne.
La fibra nervosa termina congiungendosi al sarcolemma attraverso la sinapsi, che appunto è la componente terminale dell’ assone.
Tra la membrana della sinapsi e la membrana muscolare vi è uno spazio, la fessura sinaptica, in cui sono localizzati gli ioni sodio.
All’interno della sinapsi si trovano delle vescicole contenenti il neurotrasmettitore acetilcolina, che attraversano la membrana sinaptica raggiunti dal potenziale d’azione, vanno a depositarsi sui canali del sodio trans-sarcolemmali determinandone l’apertura.
In questo modo il sodio passa attraverso il canale, generando un cambiamento di carica che provoca la depolarizzazione della membrana della fibra muscolare.
Questo fenomeno consente all’impulso nervoso di propagarsi lungo i tubuli T del reticolo, stimolando le cisterne a rilasciare ioni calcio (fino a quel momento tenuti dentro le cisterne grazie a pompe ad attività ATPasica), che a loro volta attraverso il reticolo vengono rilasciati nel sarcoplasma (citosol della fibra muscolare).
VIAGGIO ALL’ INTERNO DELLA FIBRA MUSCOLARE
La fibra muscolare è un tipo di cellula molto particolare. Possiede una forma allungata ed in ogni fibra sono presenti più nuclei. In alcuni muscoli, come quelli della coscia, le fibre possono essere lunghe fino a trenta centimetri.
Questa particolare caratteristica consente ad una sola fibra muscolare di estendersi per tutta la lunghezza del muscolo.
Tuttavia le fibre muscolari hanno un diametro che va da dieci a cento micrometri. Per intenderci cento micrometri corrispondono ad un decimo di millimetro.
Un muscolo di grandezza media contiene circa 250 000 fibre muscolari.
La miofibrilla è l’unità contrattile della cellula muscolare.
All’interno di una fibra si susseguono centinaia di miofibrille, a loro volta composte da decine di migliaia di sarcomeri contigui collegati tra loro uno dopo l’altro.
Un sarcomero è composto dai miofilamenti di actina e miosina, oltre ad altre proteine strutturali che assicurano allineamento e ancoraggio tra i miofilamenti ed il sarcolemma.
Oltre alle proteine strutturali, all’ interno della miofibrilla sono presenti due proteine fondamentali per il processo di contrazione muscolare. Queste proteine sono la troponina e la troponiosina.
I miofilamenti di actina sono composti da proteine a forma di sfera, che si susseguono a formare un polimero, che appunto è un insieme di singole unità che si susseguono.
Due di questi polimeri si intrecciano dando luogo al filamento di actina.
Ogni filamento è composto da 300-400 unità (sfere).
I filamenti di actina decorrono parallelamente tra loro e sono collegati alle estremità a formare il disco Z, a cui si connettono anche le estremità dei filamenti di actina del sarcomero contiguo.
La miosina è il miofilamento più spesso, responsabile dello scorrimento dei filamenti di actina verso il centro del sarcomero e del conseguente accorciamento della fibra muscolare.
È costituita da più subunità. Le subunità maggiori si compongono di una porzione orientata verso il sarcomero, la coda, e le teste, orientare all’estremità e rivolte verso i siti di legame dei filamenti di actina, grazie alla loro attività ATPasica.
Le subunità minori collegano le teste al collo.
Un miofilamento spesso è formato da 300-400 molecole di miosina.
Verso il centro del sarcomero le code sono unite da diverse proteine, a formare la linea M. Tra queste vi è l’enzima creatinfosfochinasi, indispensabile per la contrazione.
Avvolta intorno ai filamenti di actina, si trova la toropomiosina.
Questa proteina ha la funzione di chiudere i siti di legame dell’actina, evitando così che le teste di miosina possano agganciarvisi, mantenendo la condizione naturale di riposo del muscolo.
La troponina, è una proteina costituita da tre siti.
Un sito mantiene lo stato di riposo, inibendo il legame con la tropomiosina.
Un altro sito è invece affine con la tropomiosina. Il terzo sito è affine agli ioni calcio.
Gli ioni calcio spingono il sito della troponina affine alla tropomiosina a legarvisi, liberando i siti attivi, che divengono a loro volta sito di legame per le teste di miosina.
Quando il calcio lega il suo sito specifico sulla molecola di troponina, la spinge sulla tropomoosina che a sua volta libera i siti attivi per le teste della miosina.
Le teste della miosina ruotano verso il centro del sarcomero traendo energia dall’idrolisi di una molecola di ATP, provocando lo scorrimento dei filamenti di actina ed il conseguente accorciamento del sarcomero.
Le teste di miosina rimangono sui siti attivi dell’actina fino a quando una molecola di ATP non si lega alla testa, provocandone il distacco e l’inattivazione del sito, fino ad un successivo stimolo (gli stimoli sono 50-100 al secondo).
CONTRAZIONE MUSCOLARE, CONNUBIO TRQ IMPULSO NERVOSO E METABOLISMO CELLULARE
Il ciclo della contrazione muscolare può riprendere fintanto che vi è calcio a sufficienza per liberare i siti attivi sulle molecole di actina, che dipende dal crescente accumulo nell’ambiente contrattile del muscolo di ioni idrogeno liberati dall’ ossidazione dell’acido lattico a lattato, che è la vera causa dell’acidosi.
Il lattato funge da tampone fino a quando la concentrazione di ioni idrogeno non supera la concentrazione di lattato, provocando l’arresto della glicolisi anaerobica ed il rilascio di calcio.
Questo fenomeno viene definito cedimento metabolico, ma avviene con l’utilizzo di resistenze inquadrate all’interno di zone d’intensità più basse, che consentono tempi sotto tensione più lunghi.
Il cedimento metabolico conduce all’ipertrofia, ma attraverso meccanismi diversi, che non riguardano il solo stress meccanico.
Riducendo la resistenza è possibile proseguire lo sforzo, ma si verificherebbe un crollo delle frequenze di reclutamento.
È proprio la frequenza dell’impulso a determinare la resistenza che è possibile utilizzare.
La fibra nervosa può non condurre alcun impulso oppure condurne fino a mille al secondo, obbedendo alla legge del tutto o nulla.
La frequenza degli impulsi rappresenta il linguaggio delle fibre nervose. Dalla differenza di frequenza dipende l’effetto che la fibra nervosa ha sul tessuto innervato.
Questo significa che siccome è necessario almeno un impulso nervoso in un movimento qualunque come può essere quello di flessione del gomito, nel conseguente reclutamento di un’intera catena cinetica (gruppi muscolari), l’entità del reclutamento dipende dalla quantità di impulsi condotti nell’unità di tempo.
La frequenza determina se l’oggetto da sollevare sarà una forchetta o un bilanciere da 50 chilogrammi in un curl con bilanciere.
Di Scilipoti Nino